
The flowers on one of these plants conceal drops of sticky nectar. The other is a cheating orchid, presenting empty flowers and false promises. Can you tell which is which? Even if you knew which one carried nectar, how can you tell the difference between them? The two plants might look a bit different to human high-res optics, but now try blurring your eyes. Pretty similar, huh?
What about this pair?
 If it’s difficult for our brains and eyes to discern the difference between the flower with the reward and the one that’s falsely advertising, then what hope does a nectar-hunting fly with low resolution compound eyes and a smear of a central nervous system have?
If it’s difficult for our brains and eyes to discern the difference between the flower with the reward and the one that’s falsely advertising, then what hope does a nectar-hunting fly with low resolution compound eyes and a smear of a central nervous system have?
Specifically, I’m talking about this fly…

If this fly looks embarrassed, its because it has orchid pollen stuck to its face.
Until now, you probably thought lion, or elephant, or rhino were the most impressive animals roaming the grasslands of southern Africa. Well you’re wrong, and it’s ok to change your mind after seeing the majestic long-proboscid fly of South Africa. There are several species of these magnificent beasts, and this one is named Prosoeca ganglbaueri.

That giant proboscis hanging from its face is a tool crafted by evolution for sucking nectar from the bottom of long flower tubes, and it can grow as long as 5 cm (which is longer than the fly’s own body length). Unlike butterflies who coil their proboscises, the long-proboscid flies simply hinge the instrument down, tucking it away underneath their bodies to trail out behind them. And this species isn’t even the most extreme: proboscises in Moegistorhynchus longirostris get up to 8 cm!
Sometimes handling that long instrument can be a challenge…

In some areas of South Africa, P. ganglbaueri is the only creature capable of extracting nectar from flowers with very long floral tubes, and because of this it has become the exclusive pollinator for 20 species of plant. Altogether, the long-proboscid flies as a group bear the great responsibility as the only pollinator for approximately 130 species of plant, making them a truly important creature for the ongoing survival of many South African plants.

Figure 1 from Whitehead et al. (2018): Prosoeca ganglbaueri feeding from a variety of nectar sources. (a) Zaluzianskya microsiphon, (b) Scabiosa columbaria, (c) Agapanthus campanulatus, (d) Dianthus basuticus.
An interesting fact about flowers that are pollinated by long-proboscid flies, is that most of them are pink, or white, or some variation in between (with one blue exception). This strong colour preference is a critical feature directing the evolution of the cheating orchid flowers introduced earlier. For a deceptive orchid to attract this fly, the orchids’ flower colour must match the flies’ colour preference, or the mimicry simply won’t work.
In my recent paper, we asked whether the colour preference of flies was something that they learned, like we learn to associate that perfect golden-brown hue of fried food with a mouth-watering culinary experience, or if it was instead a more hardwired innate response, like a moth drawn to a lamp. The answer is important for understanding ultimately what is driving the evolution of false advertisement signals in mimic orchids. So, for example, if flies had an innate bias to pink or white, then cheating orchid flowers would evolve to match that bias, in the same way that any good advertisements are designed to appeal to the fundamental desires of its audience. On the other hand, if flies learned to associate nectar reward with certain colours, their preference should be determined by the colour of their local nectar diet. Under the learned scenario, orchids should be evolving to match local flowers’ colours, not any intrinsic bias of the fly.
To test this, I took advantage of just how easy it is to bamboozle these flies. With a home-made artificial flower, painted to match the pink and white flowers visited by the fly, anyone can fool a fly into attempting to feed. So I mounted a pink and a white model to my “interview stick”, and travelled across the rugged Drakensberg Mountains to interview various populations of flies. In each location, I recorded whether the local flies preferred probing the pink or white model flower, as well as the colour and species of flower that the flies were using for nectar there.
The results were clear. Flies used to feeding mainly on pink flowers preferred the pink model. Flies that fed mainly on white flowers preferred the white model. And flies that fed on both pink, white, and violet flowers, showed no clear preference between pink and white.
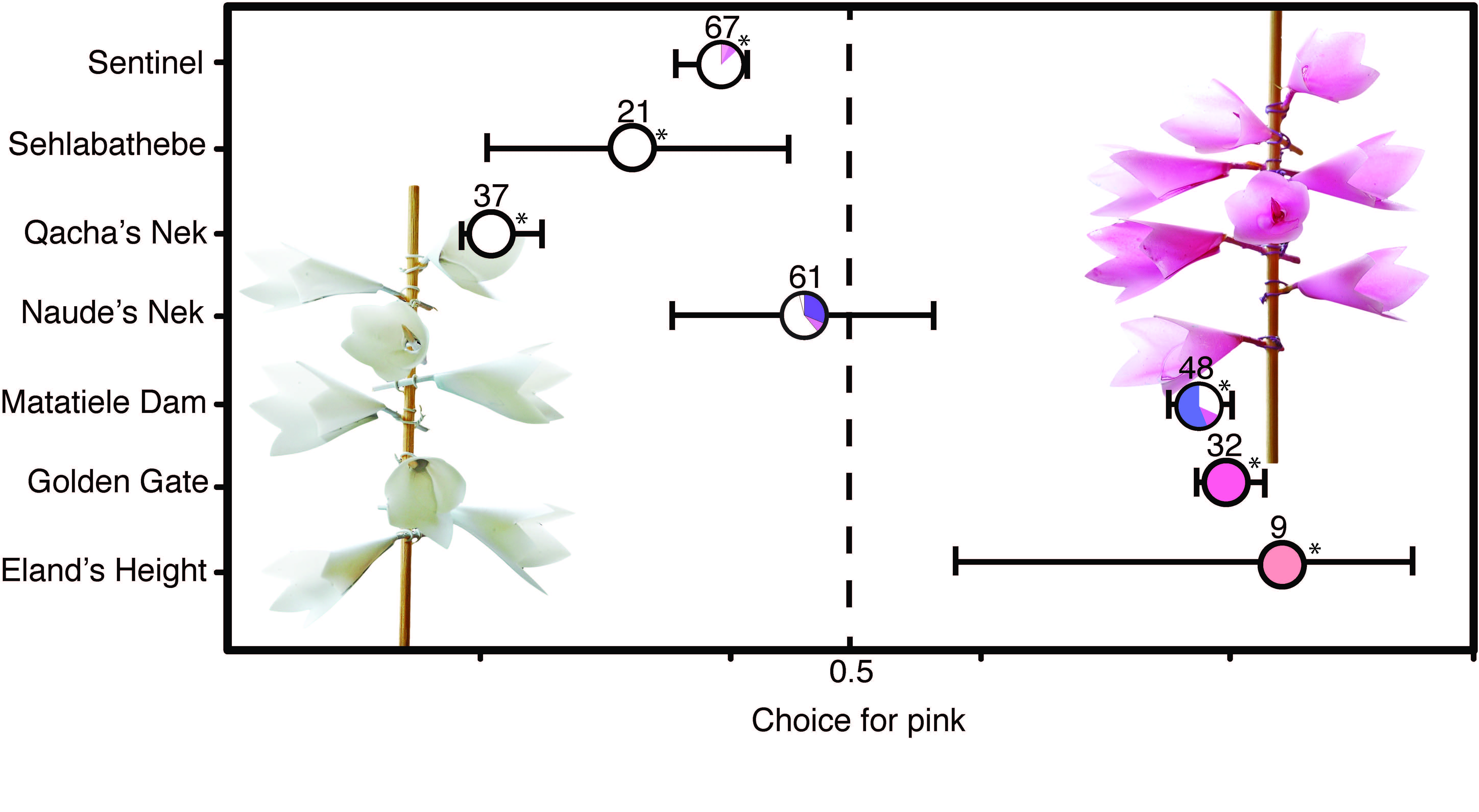
Figure 3 from Whitehead et al (2018): Pink-white preference for flies at seven sites. The x-axis shows colour preference, with pink on the right, white on the left. Measured preference at seven sites is represented, with the colour of local nectar sources depicted in the small pie charts.
This tells us that the flies are very flexible in their preferences, and the strong implication is that these flies are learning to associate colour and reward. A further result showed that as the variation of colours flies fed from increased, this made them less choosy in the pink-white preference choice. So the bottom line is that the colour of their local nectar-buffet strongly controls a fly’s colour preference.
What does this mean for orchid cheats? Well, the colour of nectar cheats is all important, and what matters most for the success of a deceptive orchid is the colour composition of the surrounding nectar-rich floral community.

Post-script:
Still wondering about which flowers in the opening images were cheats, and which had nectar?
In both cases the deceptive orchid is on the left. The first image features Disa nivea (left), and Zaluzianskya microsiphon (right), the second features Disa pulchra (left) and Watsonia lepida (right).
Reference:
Whitehead MR, Gaskett AC, Johnson SD. (in press) Floral community predicts pollinators’ color preference: implications for Batesian floral mimicry. Behavioral Ecology
































































